If evolution had a favorite body plan, it might just be the crab.
This idea lies at the heart of carcinization, which is the repeated, independent evolution of crab-like body forms across diverse crustacean lineages. The term, first coined by zoologist Lancelot Borradaile in 1916 as “one of the many attempts of Nature to evolve a crab,” carcinization has since become one of evolutionary biology’s most fascinating examples of convergent evolution. However, as McLaughlin and Lemaitre argue in their 1997 review, Carcinization in the Anomura – fact or fiction?, the phenomenon is more complex than a linear march towards “true crabs.” More recent research, in Wolfe et al. (2021), How to Become a Crab: Phenotypic Constraints on a Recurring Body Plan, reframes carcinization as a repeated evolutionary outcome shaped not only by natural selection, but by deep developmental and structural constraints.
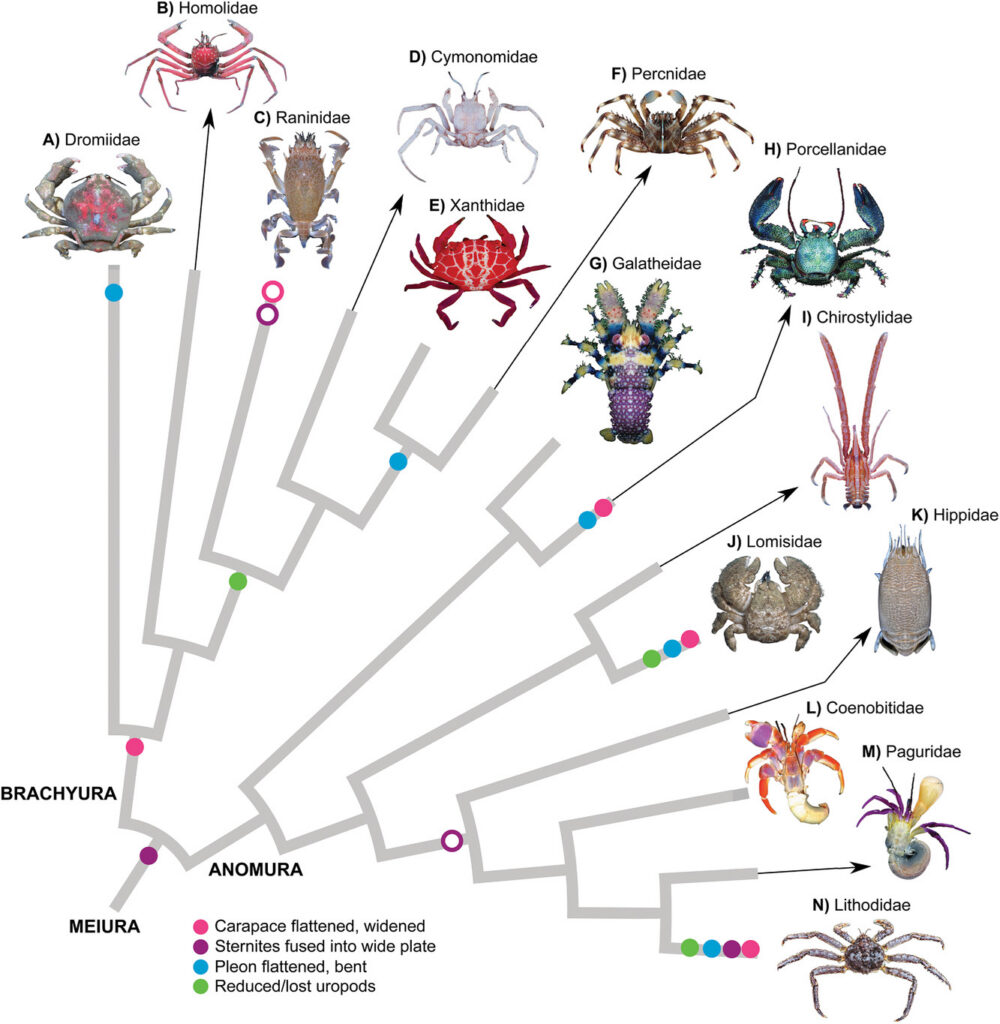
Across decapod crustaceans, crab-like forms have evolved multiple times. Within the Anomura alone (Figure 1), a group that includes hermit crabs, king crabs, squat lobsters, porcelain crabs, and coconut crabs, independent lineages have converged on broadly similar morphologies: a widened, flattened carapace, reduced and folded abdomen (pleon), and a compact body plan adapted for seafloor life. Porcelain crabs (Porcellanidae), king crabs (Lithodidae), and the terrestrial coconut crab (Birgus latro) all appear, at first glance, to have independently become crabs (McLaughlin & Lemaitre 1997).
Figure 1. Convergent evolution of crab-like body forms (carcinization) across major decapod crustacean groups, including true crabs (Brachyura), porcelain crabs (Porcellanidae), hairy stone crabs (Lomisidae), and king crabs (Lithodoidea) (Wolfe et al. 2021).
McLaughlin and Lemaitre emphasize that these similarities reflect multiple independent origins rather than a single evolutionary trajectory. Crucially, they challenge older, linear interpretations of crab evolution, such as the idea that king crabs evolved directly from hermit crabs through a gradual transformation. Instead, they argue that “carcinized” forms evolve through a variety of anatomical routes and should not be interpreted as stages in a single progression toward a crab “ideal state.”
Wolfe et al. extends this argument by reframing carcinization as part of a broader pattern of phenotypic constraint and integration. Rather than viewing crab-like forms as independently assembled traits, they propose that key features, such as carapace widening, abdominal folding, skeletal reinforcement, and locomotor reorganization, are developmentally and functionally linked. This phenotypic integration means that once certain traits evolve, they bias the evolution of others, channeling morphology toward a crab-like configuration. In this sense, carcinization is not simply selection acting on isolated traits, but the coordinated evolution of an interconnected body system (Wolfe et al. 2021).
This perspective also helps to explain why crab-like forms are so recurring. A compact, flattened body provides clear ecological advantages, including protection through a reduced exposed surface area, improved maneuverability in complex habitats, and enhanced stability for seafloor locomotion. However, neither paper supports the idea that these advantages alone guarantee convergence. Instead, Wolfe et al. emphasizes that carcinization emerges from the interaction between ecological pressures and developmental architecture that constrains how bodies can be reorganized.
Importantly, carcinization is not a one-way evolutionary trend. Both McLaughlin and Lemaitre and Wolfe et al. highlight the existence of “decarcinization”, which is the evolutionary loss of crab-like features. For example, frog crabs (Raninoidea) evolved more elongated, less compact body forms despite belonging to the true crabs, while the fossil crab Callichimaera perplexa displayed several unusual non-crab-like traits (Wolfe et al. 2021). Some lineages become more crab-like over time, while others revert toward more elongated or exposed body forms. This bidirectional pattern undermines any notion of evolutionary inevitability and suggests a changing landscape of morphological possibilities shaped by trade-offs between protection, mobility, and ecological specialization.
Fossil evidence further complicates the picture (Figure 2). Early crab-like fossils often display mosaic combinations of traits, involving partial abdominal folding, intermediate carapace shapes, or ambiguous limb structures, making it difficult to reconstruct a single pathway toward modern crabs. Wolfe et al. argue that these fossils support a stepwise and potentially repeated assembly of crab-like forms rather than a single origin of the “crab body plan.”

Figure 2. Fossil examples of crab evolution showing uncarcinized, carcinized, and decarcinized body forms across major decapod lineages. Uncarcinized forms include the more elongated, lobster-like body plans shown in A, C, and D. Carcinized forms, characterized by broadened carapaces and reduced abdomens, are shown in B and H-J. Decarcinized forms, which exhibit partial reversals away from the classic crab-like body plan, are shown in E-G. Together, these fossils illustrate how crab-like morphologies evolved repeatedly through time, while some lineages also lost or reversed key crab-like traits (Wolfe et al., 2021).
At the center of Wolfe et al. is a shift in how carcinization is interpreted. Earlier accounts sometimes framed it as evolution repeatedly “trying to make a crab,” implying a directional or goal-like process. Both McLaughlin and Lemaitre and Wolfe et al. reject this framing. Instead, they argue that crab-like morphologies repeatedly evolve because they can arise through relatively simple modifications of existing crustacean body plans, given how crustacean body plans are developmentally organized. Evolution does not aim toward crabs; rather, crab-like forms repeatedly arise because they are among the structurally available solutions to similar ecological challenges.
This has broader implications for evolution. Wolfe et al. suggests that carcinization is best understood as a case study in how constraints and integration can shape the predictability of evolution. In some contexts, evolution may appear highly repeatable, not because outcomes are predetermined, but because developmental systems channel variation along restricted and recurrent pathways. At the same time, the existence of multiple independent origins and reversals shows that these pathways are not exclusive or deterministic (Wolfe et al. 2021).
So, is carcinization happening to everybody?
Not exactly. Yes, in the sense that crab-like body plans have evolved repeatedly across independent lineages, suggesting strong functional and developmental biases toward this form. But no, because carcinization is neither universal nor inevitable. Many crustaceans never approach a crab-like form, and even within carcinized groups, evolution frequently reverses direction or produces only partial convergence (Wolfe et al. 2021, McLaughlin & Lemaitre 1997). Similar patterns of convergent evolution occur throughout nature, including the repeated evolution of streamlined, torpedo-shaped swimming bodies in sharks, dolphins, and ichthyosaurs in response to the limitations of moving through water (Futuyma & Kirkpatrick, 2017), illustrating how comparable environmental pressures can produce similar morphologies in distantly related lineages.
Ultimately, carcinization reveals something more subtle than a march toward a single optimal design. It highlights how evolution operates within a landscape shaped by ecological opportunity, developmental constraint, and historical contingency. The crab body plan is not evolution’s endpoint; it is a repeatedly accessible solution that emerges when different evolutionary paths converge on similar structural answers to similar problems.
Literature Cited
- Futuyma, D. J., & Kirkpatrick, M. (2017). Evolution (4th ed.). Sinauer Associates.
- McLaughlin, P. A., & Lemaitre, R. (1997). Carcinization in the Anomura—Fact or fiction? I. Evidence from adult morphology. Contributions to Zoology, 67(2), 79–123.
- Wolfe, J. M., Luque, J., & Bracken-Grissom, H. D. (2021). How to become a crab: Phenotypic constraints on a recurring body plan. BioEssays, 43(5), Article 2100020. https://doi.org/10.1002/bies.202100020



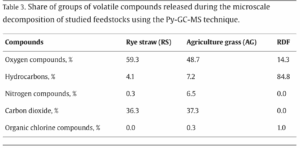
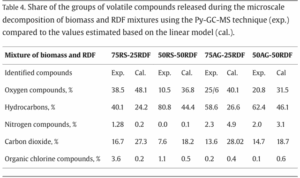
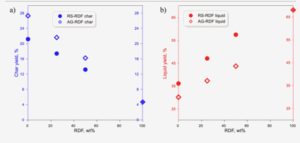
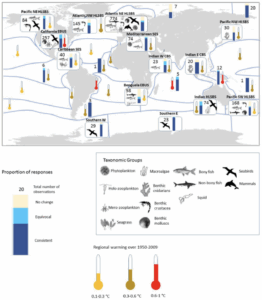












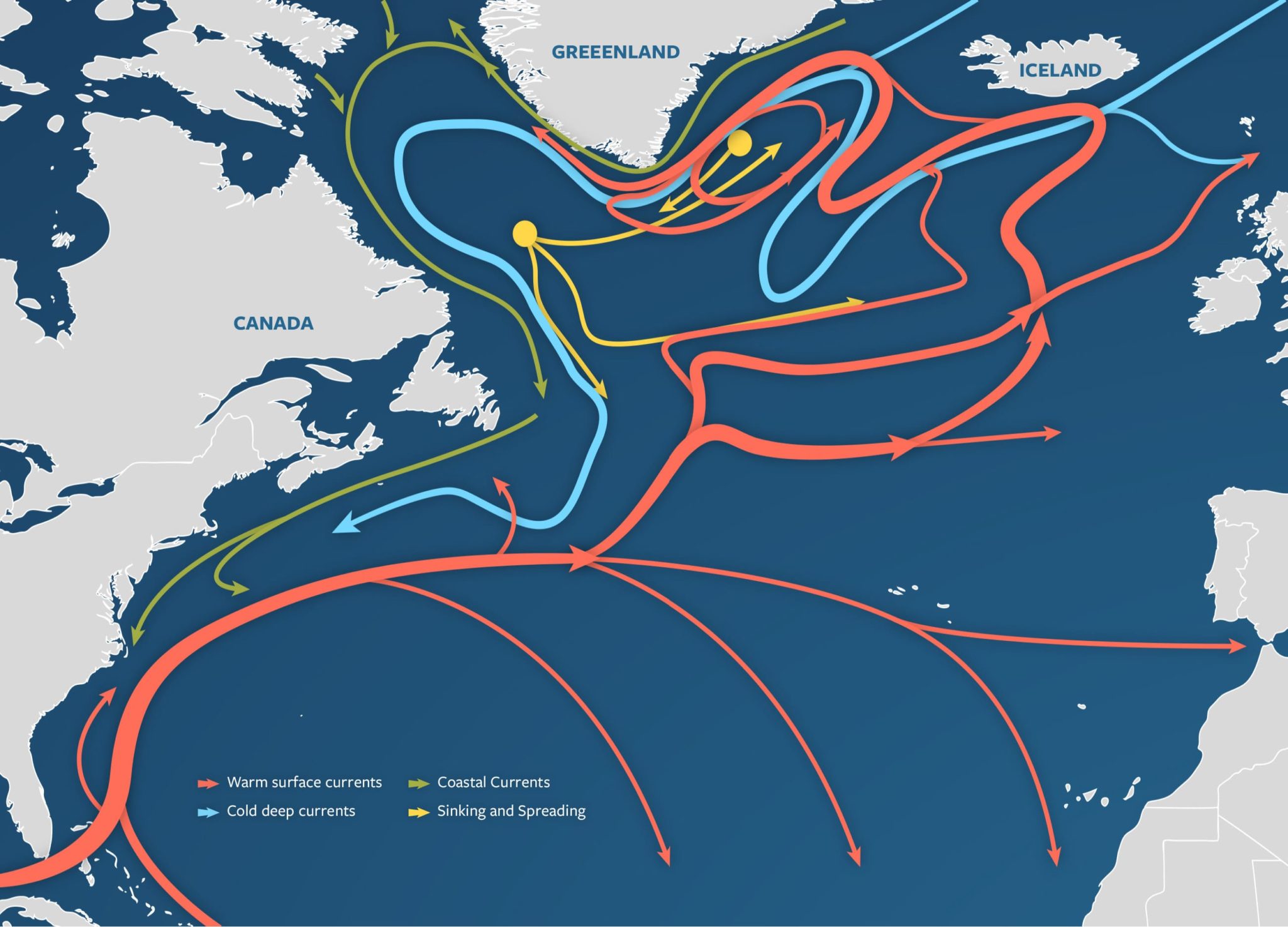 Figure 1: A visualization of the Atlantic Meridional Overturning Circulation (Adapted from “The Ocean Conveyor – Woods Hole Oceanographic Institution,” n.d.).
Figure 1: A visualization of the Atlantic Meridional Overturning Circulation (Adapted from “The Ocean Conveyor – Woods Hole Oceanographic Institution,” n.d.). Figure 2: AMOC strength at 1000m depth and 26° N latitude. Yellow band shows the range of previously observed AMOC strength (Adapted from Van Westen et al., 2024).
Figure 2: AMOC strength at 1000m depth and 26° N latitude. Yellow band shows the range of previously observed AMOC strength (Adapted from Van Westen et al., 2024).